Évolution de l'abondance des arbres indigènes communs en forêt
Évolution de l'abondance des arbres indigènes communs en forêt
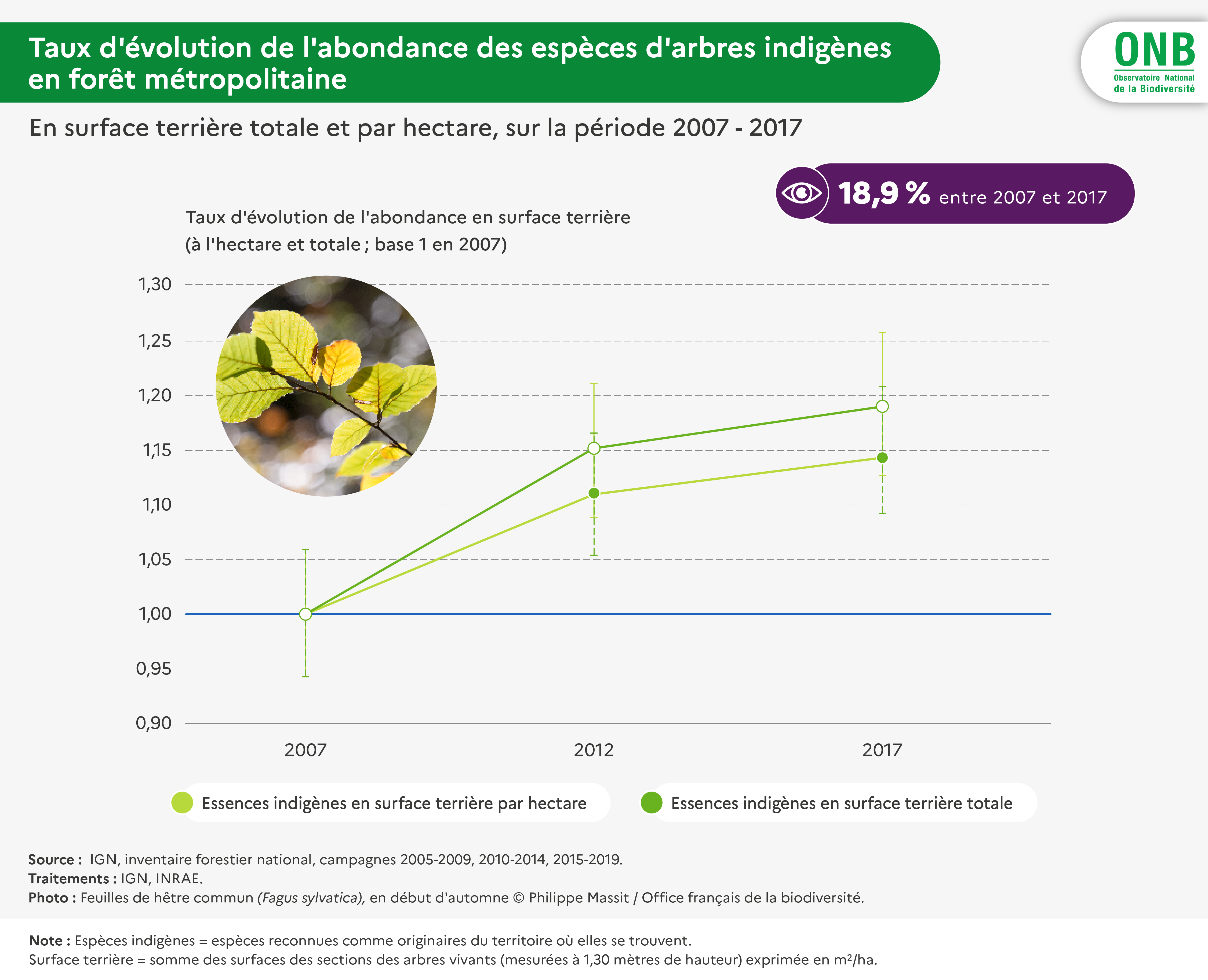
En forêt, l’abondance des espèces communes d’arbres indigènes a augmenté de 18,9% entre 2007 et 2017.
+18,9 %
en 2017
Imprimer la fiche de l'indicateur Imprimer la fiche de l'indicateur




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Définition:
Milieux concernés
Milieux forestiers
A quelle(s) question(s) répond cet indicateur
Interprétation de l'indicateur
Jeux d'indicateurs
Producteur:
Inventaire forestier de l’IGN et INRAE
Origine des données
- valeur de référence en 2007 fixée à 0% (campagnes d'inventaire 2005-2009) ;
- valeur 2012 (campagnes d'inventaire 2010-2014) ;
- valeur 2017 (campagnes d'iventaire 2015-2019).
Disponibilité des valeurs
> 5 ans
Rupture de série
Non
Méthode
La valeur de l’indicateur rencontre des problèmes quand une des espèces atteint de très faibles abondances ou disparaît : en cas d’extinction d’une espèce, la valeur de l’indicateur devient nulle. Par ailleurs, quand la moyenne est trop faible par rapport à l'écart type estimé, la distribution probabiliste suivie par l'estimateur est mal connue (Lajeunesse 2015). Pour éviter ce type de problème, il ne faut considérer que des espèces suffisamment abondantes. Après avoir essayé plusieurs méthodes, nous avons ici fait le choix de ne retenir que les espèces suffisamment abondantes pendant toutes les périodes de 5 années couvertes par l'indicateur, car définir les espèces communes sur la première période uniquement implique un biais dans la métrique (Palmer, 1993). Ainsi, seules les espèces dont le demi-intervalle de confiance à 95% était inférieur à 1.96/3 fois la valeur estimée durant toutes les périodes d’inventaire ont été considérées (valeur inspirée de Lajeunesse 2015).
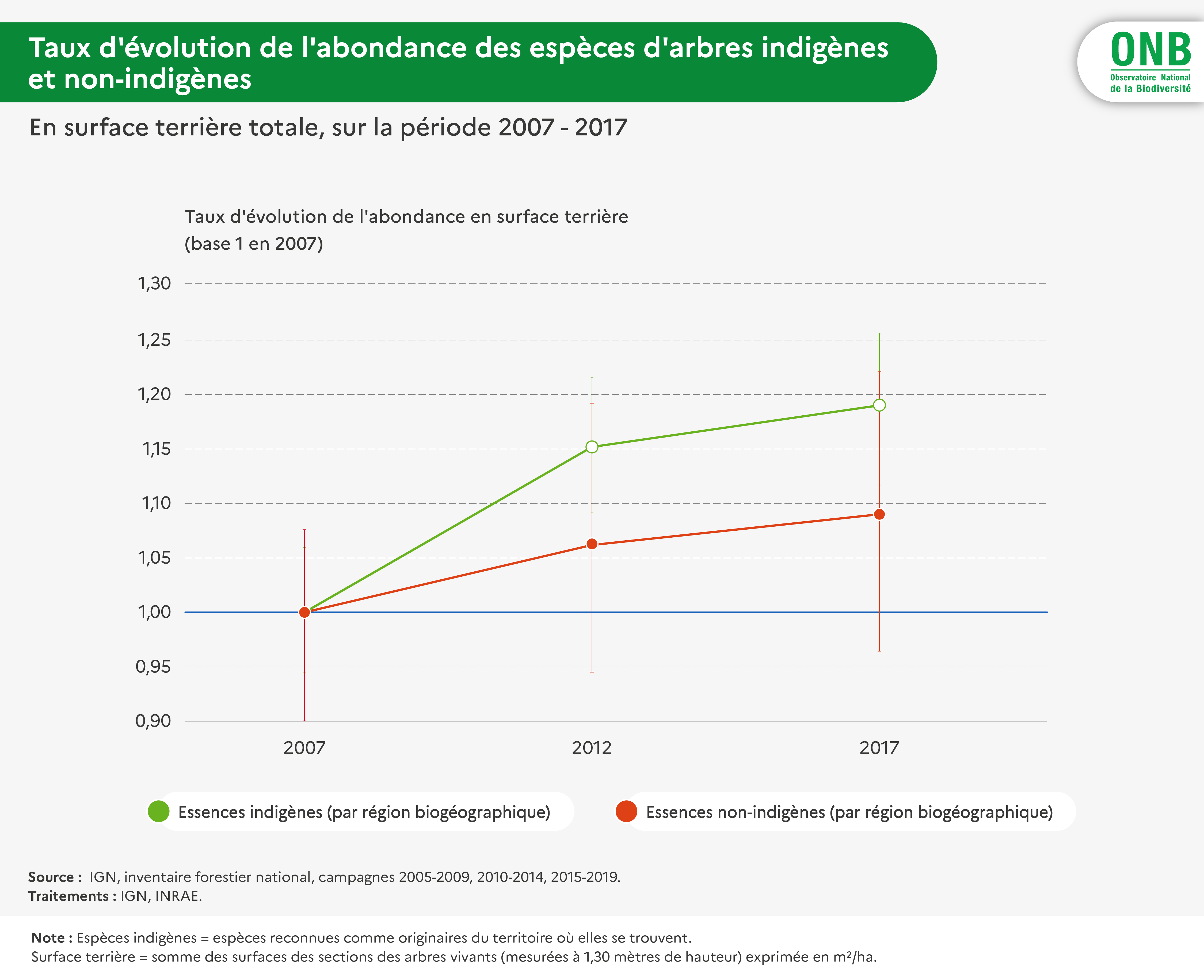
Par ailleurs, l’indicateur s’intéresse en priorité aux espèces indigènes. Le caractère indigène des espèces a été apprécié à l’échelle de la région biogéographique, à partir des listes d’indigénat établies par l’IGN pour le rapportage à l’Union européenne de l’état de conservation des habitats (édition 2013-2018). Ainsi une espèce peut être considérée indigène dans une région biogéographique et non-indigène dans une autre (ex : sapin).
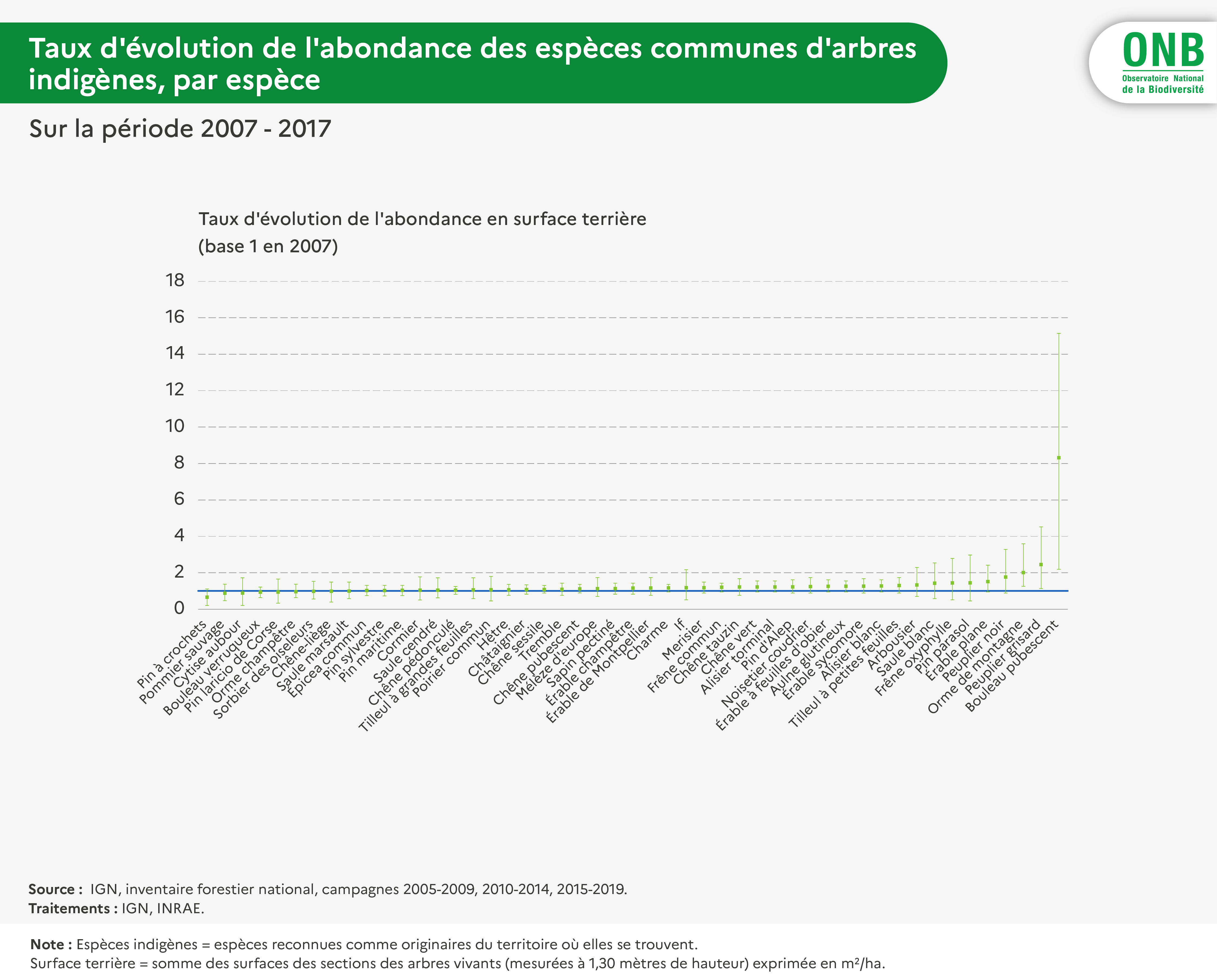
Ces sélections ont abouti à la définition d’une liste de 49 espèces indigènes communes (dont la liste se trouve dans le fichier de données disponible au téléchargement).
L’abondance est ici considérée en surface terrière, variable d’abondance des arbres utilisée très fréquemment dans le monde forestier. Elle mesure la somme totale ou par hectare des sections des troncs d’arbres à 1,3 m de hauteur. Elle est fournie par espèce et par région biogéographique, pour des périodes de cinq années par l’inventaire forestier. Elle prend la forme X ± Y m2, Y étant le demi-intervalle de confiance à 95% de chance de comprendre la valeur réelle du paramètre estimé (ici la surface terrière par espèce). À partir de ces informations, on calcule les mêmes quantités (moyenne et demi intervalle de confiance) sur la réunion des zones biogéographiques d’indigénat de l’espèce.
Calculs par espèce et par groupe d'espèces, une aide à l'interprétation de l'indicateur principal
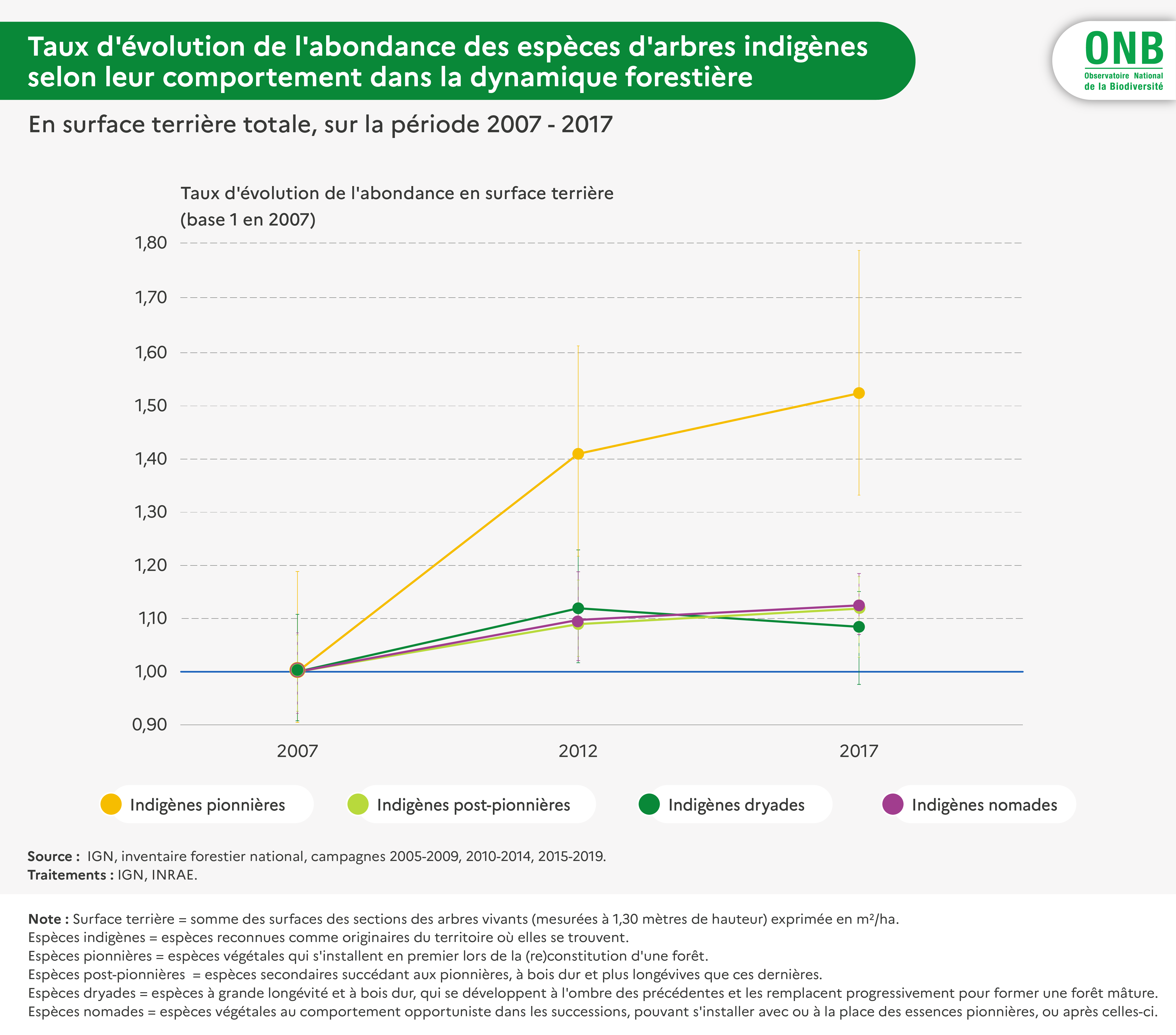
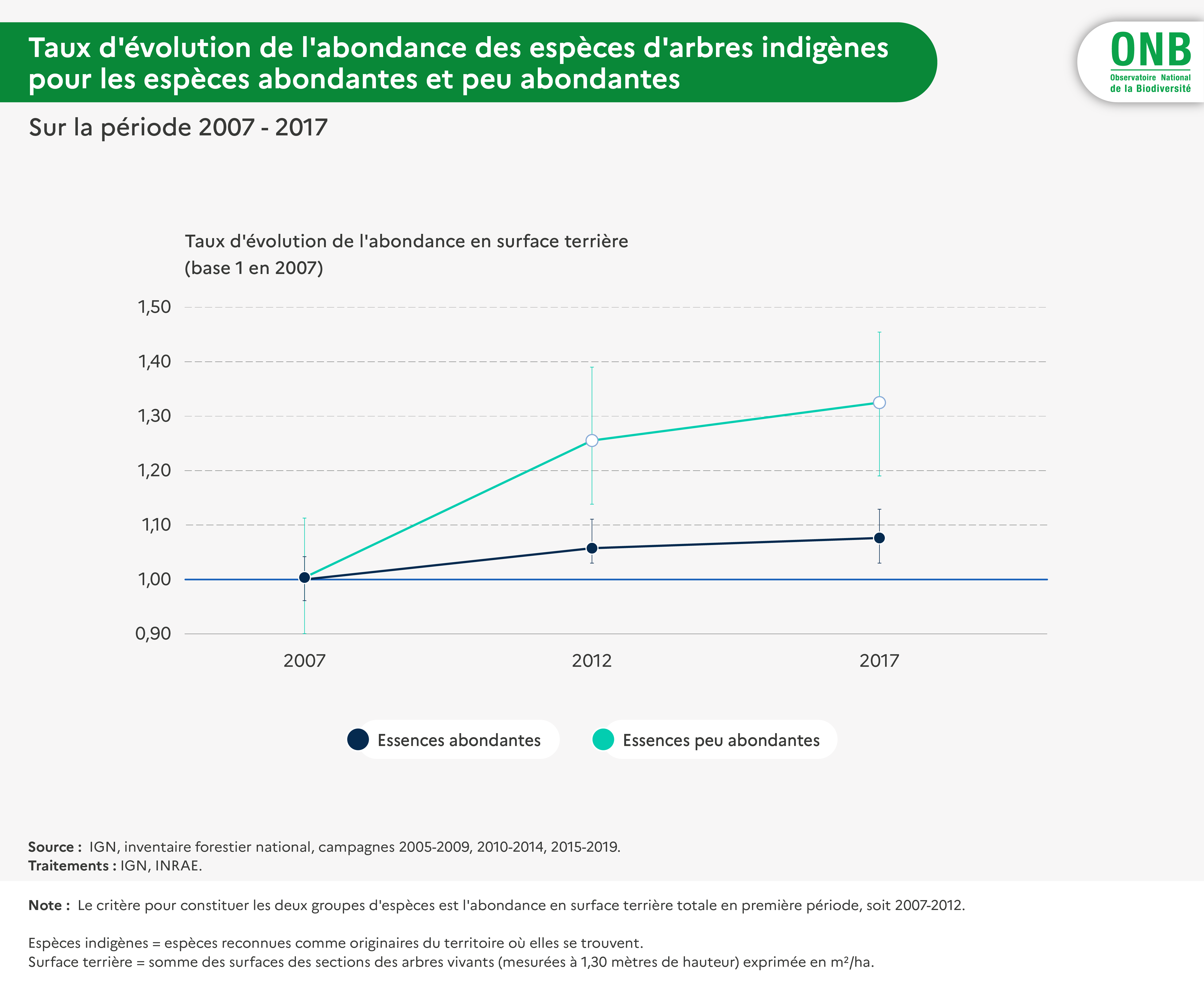
Pour interpréter l'indicateur concernant les espèces indigènes, il a été jugé intéressant de le calculer également par espèce et pour différents groupes d'espèces : espèces regroupées selon leur type biologique (Raunkier), selon leur groupe successionnel, selon leur caractère abondant ou non durant la première période (par moitiés, séparation qui se trouve correspondre à un léger décrochement d'abondance).
Par ailleurs, l'indicateur a également été calculé pour les espèces non-indigènes communes, afin de comparer sa valeur à celle obtenue pour les espèces indigènes.
Moyenne d'abondance entre-espèces (y compris par groupe d'espèces)
La moyenne géométrique de l’abondance entre espèces est calculée en :
- tirant au sort dans une loi log-normale (qui donne des valeurs strictement positives), produisant les mêmes moyenne et intervalle de confiance que celles de l’Inventaire, 10 000 valeurs aléatoires pour chaque espèce ;
- pour chacune des séries de 10 000 valeurs aléatoires et chacune des périodes, calculant la moyenne géométrique (entre espèces) des valeurs ;
- calculant ensuite le taux d’évolution de ces moyennes en divisant les 10 000 moyennes géométriques ainsi obtenues pour chaque période par la moyenne arithmétique des moyennes géométriques de la 1ère période qui est prise pour référence ;
- puis en synthétisant le ratio des 10 000 valeurs par les 10 000 valeurs (ou une permutation aléatoire de ces valeurs) correspondantes de la première période (moyenne, intervalle de confiance) ;
- transformant enfin la valeur en soustrayant 1 et en multipliant par 100 pour arriver à un pourcentage. Ainsi une valeur positive exprimera une augmentation d’abondance alors qu’une valeur négative exprimera une baisse de l’abondance moyenne.
Les chiffres d’abondance par espèce sont analysés de la même façon, excepté qu’on ne prend plus la moyenne géométrique entre espèces à la deuxième étape.
La moyenne géométrique a l’avantage de bien refléter la moyenne – sur une échelle logarithmique – des taux multiplicatifs d’évolution de chacune des espèces (cf. aussi Buckland et al., 2005 et Van Strien et al., 2012). Par exemple, si une espèce voit son abondance multipliée par 1,1 alors qu’une espèce moins abondante que la première voit son abondance divisée par 1,1, le taux d’évolution moyen basé sur la moyenne géométrique sera de 1 (soit une valeur de l’indicateur de 0%), indiquant une stabilité globale, alors que le taux de la moyenne arithmétique sera supérieur à 1 (soit une valeur de l’indicateur positive) et indiquera une augmentation de la biodiversité, et dépendant du ratio initial entre abondance de l’espèce abondante sur celle de l’espèce moins abondante.
Robustesse
Très robuste
Précision
Très précis
Sensibilité
Sensible
Efficacité
Très efficace
Accessibilité des données
Facilement accessibles
Homogénéité des données
Très homogènes
Fiabilité des données
Très fiables
Pérennité des données
Pérennité garantie
Abondance des données
Abondantes
Avantages
Il permet, à partir des évolutions par espèce qui sont calculées, d’interpréter la valeur globale de l’indicateur. Il est déclinable par groupe d’espèces d’intérêt afin de pouvoir interpréter les résultats globaux.
Limites
L'indicateur n’est pas adapté pour suivre les espèces les plus rares par définition.
Piste d'améliorations
(ii) Calculer l’indicateur à l’échelle de la région biogéographique, et calculer l'indicateur avec des données de densité et de volume en plus de celles de surface terrière.
(iii) Remonter plus loin dans le temps à partir des données anciennes de l’inventaire forestier. On perdrait alors en précision sur la définition des espèces car avant 2005, les données des arbres peuvent assembler en une seule essence plusieurs espèces (par ex. une seule essence de bouleau pour les bouleaux pubescent et verruqueux). Pour savoir si cela apporterait des informations intéressantes, il faudrait le calculer. Pour la première édition, on se limite à un retour à 2005 en conservant la précision sur 49 espèces communes d’arbres. (iv) Renseigner l'indigénat par SylvoEcoRégion plutôt que par Région Biogéographique pour pallier à certains problèmes de qualification de l'indigénat.
(v) Envisager le calcul de l’indicateur sur la base de données annuelles d’abondance, permettant une mise à jour plus fréquente de l’indicateur ou bien rester sur une moyenne quinquennale mais mise à jour tous les ans, de façon glissante.
(vi) Proposer d’autres métriques que la moyenne géométrique gardant des propriétés intéressantes de celle-ci tout en permettant d’incorporer toutes les espèces, y compris les espèces rares.
(vii) Tester différentes méthodes de définition des espèces communes suivant la ou les périodes utilisée(s).